Хромосомы - самовоспроизводящиеся структуры клеточного ядра. Как у прокариотических, так и у эукариотических организмов гены располагаются группами на отдельных молекулах ДНК, которые при участии белков и других макромолекул клеток организуются в хромосомы. Зрелые клетки зародышевой линии (гаметы - яйцеклетки, спермии) многоклеточных организмов содержат по одному (гаплоидному) набору хромосом организма.
После того как к полюсам отойдут полные наборы хроматид, их называют хромосомами . Хромосомы - это структуры в ядре клеток эукариот, которые пространственно и функционально организовывают ДНК в геноме индивидуумов.
Химический состав хромосом. Хромосома представляет собой дезоксирибонуклеопротеид (ДНП), то есть комплекс, образованный из одной непрерывной двухцепочечной молекулы ДНК и белков (гистонов и негистонов). В состав хромосом входят также липиды и минеральные вещества (например, ионы Ca 2+ , Mg 2+).
Каждая хромосома – сложное надмолекулярное образование , сформированное в результате компактизации хроматина.
Строение хромосом. В большинстве случаев хромосомы хорошо видны лишь в делящихся клетках начиная со стадии метафазы, когда их можно видеть даже в световой микроскоп. В этот период удается определить количество хромосом в ядре, их размеры, форму и строение. Именно такие хромосомы называют метафазными. Интерфазные хромосомы часто называют просто хроматином .
Число хромосом обычно постоянно для всех клеток особи любого вида растений, животных и человека. Но у разных видов количество хромосом неодинаково (от двух до нескольких сотен). Наименьшее число хромосом имеет лошадиная аскарида, наибольшее встречается у простейших и папоротников, для которых характерны высокие уровни полиплоидии. Обычно диплоидные наборы содержат от одного до нескольких десятков хромосом.
Количество хромосом в ядре не связано с уровнем эволюционного развития живых организмов. У многих примитивных форм оно велико, например, в ядрах некоторых видов простейших содержатся сотни хромосом, тогда как у шимпанзе их всего только 48.
Каждая хромосома, образованная одной молекулой ДНК, представляет собой удлиненную палочковидную структуру – хроматиду , имеющую два «плеча», разделенных первичной перетяжкой, или центромерой. Метафазная хромосома состоит из двух соединенных центромерой сестринских хроматид, каждая из которых содержит одну молекулу ДНК, уложенную в виде спирали.
Центромера – это небольшое фибриллярное тельце, осуществляющее первичную перетяжку хромосомы. Она является важнейшей частью хромосомы, так как определяет ее движение. Центромеру, к которой прикрепляются нити веретена во время деления (при митозе и мейозе), называют кинетохором (от греч. kinetos – подвижный и choros – место). Он контролирует движение расходящихся хромосом при делении клетки. Хромосома, лишенная центромеры, не способна совершать упорядоченное движение и может потеряться.
Обычно центромера хромосомы занимает определенное место, и это является одним из видовых признаков, по которому различают хромосомы. Изменение положения центромеры в той или иной хромосоме служит показателем хромосомных перестроек. Плечи хромосом оканчиваются участками, не способными соединяться с другими хромосомами или их фрагментами. Эти концевые участки хромосом называют теломерами . Теломеры предохраняют концы хромосом от слипания и тем самым обеспечивают сохранение их целостности. За открытие механизма защиты хромосом теломерами и ферментом теломеразой американские ученые Э. Блекберн, К. Грейдер и Д. Шостак в 2009 году были удостоены Нобелевской премии в области медицины и физиологии. Концы хромосом нередко обогащены гетерохроматином.

В зависимости от расположения центромеры определяют три основных вида хромосом: равноплечие (плечи равной длины), неравноплечие (с плечами разной длины) и палочковидные (с одним, очень длинным и другим, очень коротким, едва заметным плечом). Некоторые хромосомы имеют не только одну центромеру, но еще и вторичную перетяжку, не связанную с прикреплением нити веретена при делении. Этот участок – ядрышковый организатор , выполняющий функцию синтеза ядрышка в ядре.
Репликация хромосом
Важным свойством хромосом является их способность к удвоению (самовоспроизведению). Обычно удвоение хромосом предшествует делению клетки. В основе удвоения хромосом лежит процесс репликации (от лат. replicatio – повторение) макромолекул ДНК, обеспечивающий точное копирование генетической информации и передачу ее от поколения к поколению. Удвоение хромосом – это сложный процесс, включающий в себя не только репликацию гигантских молекул ДНК, но также синтез связанных с ДНК хромосомных белков. Конечным этапом является упаковка ДНК и белков в особые комплексы, образующие хромосому. В результате репликации вместо одной материнской хромосомы появляются две идентичные ей дочерние хромосомы.
Функция хромосом заключается:
- в хранении наследственной информации. Хромосомы являются носителями генетической информации;
- передаче наследственной информации. Наследственная информация передается путем репликации молекулы ДНК;
- реализации наследственной информации. Благодаря воспроизводству того или иного типа и-РНК и, соответственно, того или иного типа белка осуществляется контроль над всеми процессами жизнедеятельности клетки и всего организма.
Таким образом, хромосомы с заключенными в них генами обусловливают непрерывный ряд воспроизведения.
Хромосомы осуществляют сложную координацию и регуляцию процессов в клетке вследствие заключенной в них генетической информации, обеспечивающей синтез первичной структуры белков-ферментов.
У каждого вида в клетках находится определенное количество хромосом. Они являются носителями генов, определяющих наследственные свойства клеток и организмов вида. Ген – это участок молекулы ДНК хромосомы, на котором синтезируются различные молекулы РНК (трансляторы генетической информации).
В соматических, то есть телесных, клетках обычно содержится двойной, или диплоидный, набор хромосом. Он состоит из пар (2n) практически одинаковых по форме и размеру хромосом. Такие парные, похожие друг на друга хромосомные наборы называют гомологичными (от греч. homos – равный, одинаковый, общий). Они происходят от двух организмов; один набор от материнского, а другой – от отцовского. В таком парном наборе хромосом заключена вся генетическая информация клетки и организма (особи). Гомологичные хромосомы одинаковы по форме, длине, строению, расположению центромеры и несут одни и те же гены, имеющие одинаковую локализацию. Они содержат одинаковый набор генов, хотя и могут различаться их аллелями. Таким образом, гомологичные хромосомы содержат очень близкую, но не идентичную наследственную информацию.
Совокупность признаков хромосом (их число, размеры, форма и детали микроскопического строения) в клетках тела организма того или иного вида называют кариотипом. Форма хромосом, их число, размеры, расположение центромеры, наличие вторичных перетяжек всегда специфичны для каждого вида, по ним можно сопоставлять родство организмов и устанавливать их принадлежность к тому или иному виду.
Постоянство кариотипа, свойственное каждому виду, выработалось в процессе его эволюции и обусловлено закономерностями митоза и мейоза. Однако в процессе существования вида в его кариотипе вследствие мутаций могут произойти изменения хромосом. Некоторые мутации существенно изменяют наследственные качества клетки и организма в целом.
Постоянные характеристики хромосомного набора – количество и морфологические особенности хромосом, определяемые главным образом расположением центромер, наличием вторичных перетяжек, чередованием эухроматиновых и гетерохроматиновых участков и пр., позволяют идентифицировать виды. Поэтому кариотип называют «паспортом» вида .
История открытия хромосом
Рисунок из книги В. Флемминга, изображающий разные стадии деления клеток эпителия саламандры (W. Flemming. Zellsubstanz, Kern und Zelltheilung. 1882 г.)
В разных статьях и книгах приоритет открытия хромосом отдают разным людям, но чаще всего годом открытия хромосом называют 1882 год, а их первооткрывателем - немецкого анатома В. Флеминга . Однако справедливее было бы сказать, что он не открыл хромосомы, а в своей фундаментальной книге "Zellsubstanz, Kern und Zelltheilung" (нем.) собрал и упорядочил сведения о них, дополнив результатами собственных исследований. Термин «хромосома» был предложен немецким гистологом Генрихом Вальдейером в 1888 году, «хромосома» в буквальном переводе означает «окрашенное тело», поскольку оснóвные красители хорошо связываются хромосомами.
Сейчас сложно сказать, кто сделал первое описание и рисунок хромосом. В 1872 году швейцарский ботаник Карл фон Нэгили опубликовал работу, в которой изобразил некие тельца, возникающие на месте ядра во время деления клетки при образовании пыльцы у лилии (Lilium tigrinum ) и традесканции (Tradescantia ). Однако его рисунки не позволяют однозначно утверждать, что К. Нэгили видел именно хромосомы. В том же 1872 году ботаник Э. Руссов привёл свои изображения деления клеток при образовании спор у папоротника из рода ужовник (Ophioglossum ) и пыльцы лилии (Lilium bulbiferum ). На его иллюстрациях легко узнать отдельные хромосомы и стадии деления. Некоторые же исследователи полагают, что первыми увидел хромосомы немецкий ботаник Вильгельм Гофмайстер задолго до К. Нэгили и Э. Руссова, ещё в 1848-1849 годах. При этом ни К. Нэгили, ни Э. Руссов, ни тем более В. Гофмейстер не осознавали значения того, что видели.
После переоткрытия в 1900 году законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902 году Т. Бовери и в 1902-1903 годах У. Сеттон (Walter Sutton ) независимо друг от друга первыми выдвинули гипотезу о генетической роли хромосом. Т. Бовери обнаружил, что зародыш морского ежа Paracentrotus lividus может нормально развиваться только при наличии хотя бы одного, но полного набора хромосом. Также он установил, что разные хромосомы не идентичны по своему составу. У. Сеттон изучал гаметогенез у саранчового Brachystola magna и понял, что поведение хромосом в мейозе и при оплодотворении полностью объясняет закономерности расхождения менделевских факторов и образования их новых комбинаций.
Экспериментальное подтверждение этих идей и окончательное формулирование хромосомной теории было сделано в первой четверти XX века основателями классической генетики, работавшими в США с плодовой мушкой (D.melanogaster ): Т. Морганом , К. Бриджесом (C.B.Bridges ), А. Стёртевантом (A.H.Sturtevant ) и Г. Мёллером . На основе своих данных они сформулировали «хромосомную теорию наследственности», согласно которой передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Эти выводы были опубликованы в 1915 году в книге «The mechanisms of mendelian heredity» (англ.).
В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине .
Хромосомы эукариот
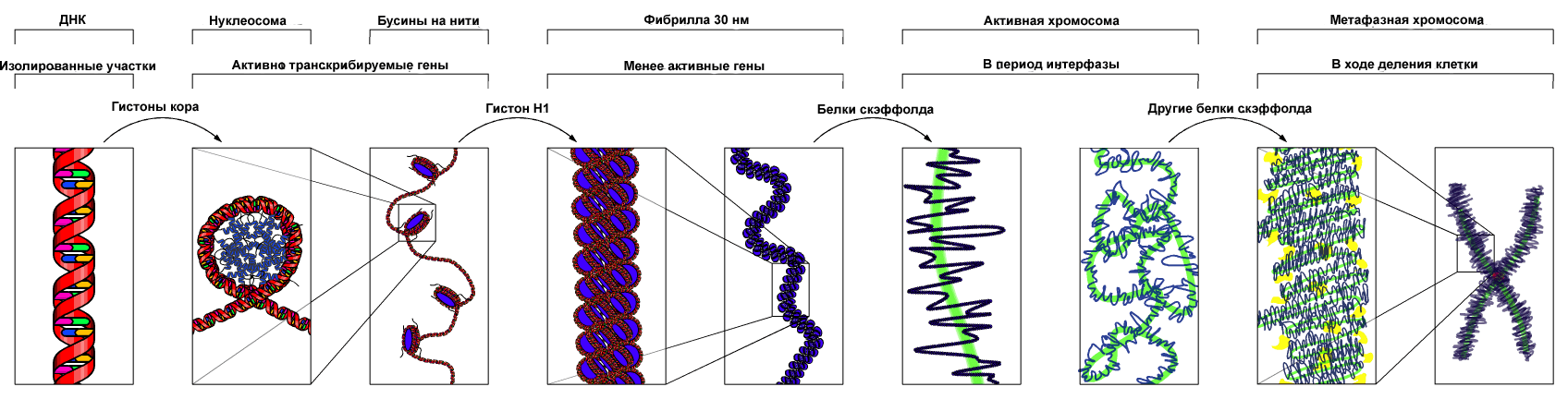
Основу хромосомы составляет линейная (не замкнутая в кольцо) макромолекула дезоксирибонуклеиновой кислоты (ДНК) значительной длины (например, в молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований). В растянутом виде длина хромосомы человека может достигать 5 см. Помимо неё, в состав хромосомы входят пять специализированных белков - H1, H2A, H2B, H3 и H4 (так называемые гистоны) и ряд негистоновых белков. Последовательность аминокислот гистонов высококонсервативна и практически не различается в самых разных группах организмов.
Первичная перетяжка
Хромосомная перетяжка (X. п.), в которой локализуется центромера и которая делит хромосому на плечи.
Вторичные перетяжки
Морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. У человека это 9, 13, 14, 15, 21 и 22 хромосомы.
Типы строения хромосом
Различают четыре типа строения хромосом:
- телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
- акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
- субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
- метацентрические (V-образные хромосомы, обладающие плечами равной длины).
Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода .
Спутники (сателлиты)
Сателлит - это округлое или удлинённое тельце, отделённое от основной части хромосомы тонкой хроматиновой нитью, по диаметру равный или несколько меньший хромосоме. Хромосомы, обладающие спутником принято обозначать SAT-хромосомами. Форма, величина спутника и связывающей его нити постоянны для каждой хромосомы.
Зона ядрышка
Зоны ядрышка (организаторы ядрышка ) - специальные участки, с которыми связано появление некоторых вторичных перетяжек.
Хромонема
Хромонема - это спиральная структура, которую удаётся увидеть в декомпактизованных хромосомах через электронный микроскоп. Впервые наблюдалась Баранецким в 1880 году в хромосомах клеток пыльников традесканции , термин ввёл Вейдовский. Хромонема может состоять из двух, четырёх и более нитей, в зависимости от исследуемого объекта. Эти нити образуют спирали двух типов:
- паранемическую (элементы спирали легко разъединить);
- плектонемическую (нити плотно переплетаются).
Хромосомные перестройки
Нарушение структуры хромосом происходит в результате спонтанных или спровоцированных изменений (например, после облучения).
- Генные (точковые) мутации (изменения на молекулярном уровне);
- Аберрации (микроскопические изменения, различимые при помощи светового микроскопа):
Гигантские хромосомы
Такие хромосомы, для которых характерны огромные размеры, можно наблюдать в некоторых клетках на определённых стадиях клеточного цикла . Например, они обнаруживаются в клетках некоторых тканей личинок двукрылых насекомых (политенные хромосомы) и в ооцитах различных позвоночных и беспозвоночных (хромосомы типа ламповых щёток). Именно на препаратах гигантских хромосом удалось выявить признаки активности генов .
Политенные хромосомы
Впервые обнаружены Бальбиани в -го, однако их цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером. Содержатся в клетках слюнных желёз , кишечника , трахей , жирового тела и мальпигиевых сосудов личинок двукрылых .
Хромосомы типа ламповых щёток
Имеются данные о наличии у бактерий белков, связанных с ДНК нуклеоида , но гистонов у них не обнаружено.
Хромосомы человека
В каждой ядросодержащей соматической клетке человека содержится 23 пары линейных хромосом, а также многочисленные копии митохондриальной ДНК . В нижеприведённой таблице показано число генов и оснований в хромосомах человека.
| Хромосома | Количество генов | Всего оснований | Секвенированых оснований |
|---|---|---|---|
| 4 234 | 247 199 719 | 224 999 719 | |
| 1 491 | 242 751 149 | 237 712 649 | |
| 1 550 | 199 446 827 | 194 704 827 | |
| 446 | 191 263 063 | 187 297 063 | |
| 609 | 180 837 866 | 177 702 766 | |
| 2 281 | 170 896 993 | 167 273 993 | |
). Хроматин неоднороден, и некоторые типы такой неоднородности видны под микроскопом. Тонкая структура хроматина в интерфазном ядре, определяемая характером укладки ДНК и ее взаимодействия с белками, играет важную роль в регуляции транскрипции генов и репликации ДНК и, возможно, клеточной дифференцировки .
Последовательности нуклеотидов ДНК, которые образуют гены и служат матрицей для синтеза мРНК , распределены по всей длине хромосом (отдельные гены, разумеется, слишком малы, чтобы их можно было видеть под микроскопом). К концу XX столетия примерно для 6000 генов было установлено, на какой хромосоме и в каком участке хромосомы они находятся и каков характер их сцепления (то есть положения друг относительно друга).
Неоднородность метафазных хромосом, как уже упоминалось, можно увидеть даже при световой микроскопии. При дифференциальном окрашивании по меньшей мере в 12 хромосомах обнаружены различия в ширине некоторых полос между гомологичными хромосомами ( рис. 66.3). Такие полиморфные участки состоят из некодирующих высокоповторяющихся последовательностей ДНК.
Методы молекулярной генетики сделали возможной идентификацию огромного числа меньших по размеру и потому не выявляемых при световой микроскопии полиморфных участков ДНК. Эти участки выявляют как полиморфизм длин рестрикционных фрагментов, варьирующие по числу тандемные повторы и полиморфизм коротких тандемных повторов (моно-, ди-, три- и тетрануклеотидных). Такая изменчивость фенотипически обычно не проявляется.
Однако полиморфизм служит удобным инструментом пренатальной диагностики благодаря сцеплению определенных маркеров с мутантными генами, вызывающими заболевания (например, при миопатии Дюшенна), а также при установлении зиготности близнецов , установлении отцовства и прогнозирования отторжения трансплантата .
Трудно переоценить значение таких маркеров, особенно широко распространенных в геноме высокополиморфных коротких тандемных повторов, для картирования генома человека. В частности, они позволяют установить точный порядок и характер взаимодействия локусов, играющих важную роль в обеспечении нормального онтогенеза и клеточной дифференцировки. Это касается и тех локусов, мутации в которых приводят к наследственным заболеваниям.
Различимые под микроскопом участки на коротком плече акроцентрических аутосом ( рис. 66.1) обеспечивают синтез рРНК и образование ядрышек , поэтому их называют районами ядрышкового организатора . В метафазе они неконденсированы и не окрашиваются. Районы ядрышкового организатора примыкают к находящимся на конце короткого плеча хромосомы конденсированным участкам хроматина - спутникам. Спутники не содержат генов и являются полиморфными участками.
В небольшой части клеток удается выявить другие деконденсированные в метафазе участки, так называемые ломкие участки , где могут происходить "полные" разрывы хромосомы. Клиническое значение имеют нарушения в единственном подобном участке, расположенном на конце длинного плеча Х-хромосомы. Такие нарушения вызывают синдром ломкой Х-хромосомы .
Другие примеры специализированных районов хромосом - теломеры и центромеры .
Пока точно не установлена роль гетерохроматина , на долю которого приходится значительная часть генома человека. Гетерохроматин конденсирован в течение практически всего клеточного цикла, он неактивен и реплицируется поздно. Большинство участков конденсированы и неактивны во всех клетках (), хотя другие, например Х-хромосома, могут быть как конденсированными и неактивными, так и деконденсированными и активными ( факультативный гетерохроматин). Если из-за хромосомных аберраций гены оказываются рядом с гетерохроматином, то активность таких генов может изменяться или даже блокироваться. Поэтому проявления хромосомных аберраций , таких, как дупликации или делеции, зависят не только от затронутых локусов, но и от типа хроматина в них. Многие хромосомные аномалии, не являющиеся летальными, затрагивают неактивные или инактивируемые участки генома. Возможно, этим объясняется, что трисомии по некоторым хромосомам или моносомии по Х-хромосоме совместимы с жизнью.
Проявления хромосомной аномалии зависят также от нового расположения структурных и регуляторных генов по отношению друг к другу и к гетерохроматину.
К счастью, многие структурные особенности хромосом удается надежно обнаружить цитологическими методами. В настоящее время существует ряд методов дифференциального окрашивания хромосом ( рис. 66.1 и рис. 66.3). Расположение и ширина полос идентичны в каждой паре гомологичных хромосом, за исключением полиморфных участков, поэтому окрашивание можно использовать в клинической цитогенетике для идентификации хромосом и выявления в них структурных нарушений.