Здесь в отличие от (3.2.1) знаки (-012) и (+a2i) разные. Как и в случае конкуренции (система уравнений (2.2.1)), начало координат (1) для этой системы является особой точкой типа «неустойчивый узел». Три других возможных стационарных состояния:

Биологический смысл требует положительности величин Х у х 2. Для выражения (3.3.4) это означает, что
В случае, если коэффициент внутривидовой конкуренции хищников а ,22 = 0, условие (3.3.5) приводит к условию ai2
Возможные типы фазовых портретов для системы уравнений (3.3.1) представлены на рис. 3.2 a-в. Изоклины горизонтальных касательных представляют собой прямые
а изоклины вертикальных касательных - прямые
Из рис. 3.2 видно следующее. Система хищник -жертва (3.3.1) может иметь устойчивое положение равновесия, в котором популяция жертв полностью вымерла (х = 0) и остались только хищники (точка 2 на рис. 3.26). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х хищник Х 2 имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для хз. Особые точки (1) и (3) (рис. 3.26) являются неустойчивыми. Вторая возможность - устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы - устойчивая точка (3) (рис. 3.2а). Здесь особая точка (1) - также неустойчивый узел.
Наконец, третья возможность - устойчивое сосуществование популяций хищника и жертвы (рис. 3.2 в), стационарные численности которых выражаются формулами (3.3.4). Рассмотрим этот случай подробнее.
Предположим равенство нулю коэффициентов внутривидовой конкуренции (аи = 0, i = 1, 2). Предположим также, что хищники питаются только жертвами вида х и в отсутствие их вымирают со скоростью С2 (в (3.3.5) С2
Проведем подробное исследование этой модели, воспользовавшись обозначениями, наиболее широко принятыми в литературе. Переобо-

Рис. 3.2. Расположение главных изоклин на фазовом портрете вольтерров- ской системы хищник жертва при разном соотношении параметров: а - о» б -
С I С2 С2
1, 3 - неустойчивые, 2 - устойчивая особая точка; в -
1, 2, 3 - неустойчивые, 4 - устойчивая особая точка значим
Система хищник-жертва в этих обозначениях имеет вид:

Свойства решений системы (3.3.6) будем исследовать на фазовой плоскости N
1
ON
2
Система имеет два стационарных решения. Их легко определить, приравняв нулю правые части системы. Получим:

Отсюда стационарные решения:

Рассмотрим подробнее второе решение. Найдем первый интеграл системы (3.3.6), не содержащий t. Умножим первое уравнение на -72, второе - на -71 и результаты сложим. Получим:
Теперь разделим первое уравнение на N и умножим на € 2, а второе разделим на JV 2 и умножим на е. Результаты снова сложим:
Сравнивая (3.3.7) и (3.3.8), будем иметь:

Интегрируя, получим:

Это и есть искомый первый интеграл. Таким образом, система (3.3.6) является консервативной, поскольку имеет первый интеграл движения, величину, представляющую собой функцию переменных системы N и N 2 и не зависящую от времени. Это свойство позволяет конструировать для вольтерровских систем систему понятий, аналогичную статистической механике (см. гл. 5), где существенную роль играет величина энергии системы, неизменная во времени.
При каждом фиксированном с > 0 (что соответствует определенным начальным данным) интегралу соответствует определенная траектория на плоскости N 1 ON 2 , служащая траекторией системы (3.3.6).
Рассмотрим графический способ построения траектории, предложенный самим Вольтерра. Заметим, что правая часть формулы (3.3.9) зависит только от Д г 2, а левая - только от N. Обозначим
Из (3.3.9) следует, что между X и Y имеется пропорциональная зависимость
На рис. 3.3 изображены первые квадранты четырех систем координат XOY, NOY , N 2 OX и Д Г 10N 2 так, чтобы все они имели общее начало координат.
В левом верхнем углу (квадрант NOY) построен график функции (3.3.8), в правом нижнем (квадрант N 2 OX) - график функции Y. Первая функция имеет min при Ni = а вторая - max при N 2 = ?-
Наконец, в квадранте XOY построим прямую (3.3.12) для некоторого фиксированного С.
Отметим точку N на оси ON . Этой точке соответствует определенное значение Y(N 1), которое легко найти, проведя перпендикуляр

Рис. 3.3.
через N до пересечения с кривой (3.3.10) (см. рис. 3.3). В свою очередь, значению К(Д^) соответствует некоторая точка М на прямой Y = сХ и, следовательно, некоторое значение X(N) = Y(N)/c, которое можно найти, проведя перпендикуляры AM и MD. Найденному значению (эта точка отмечена на рисунке буквой D) соответствуют две точки Р и G на кривой (3.3.11). По этим точкам, проводя перпендикуляры, найдем сразу две точки Е" и Е ", лежащие на кривой (3.3.9). Их координаты:

Проводя перпендикуляр AM , мы пересекли кривую (3.3.10) еще в одной точке В. Этой точке соответствуют те же Р и Q на кривой (3.3.11) и те же N и Щ. Координату N этой точки можно найти, опустив перпендикуляр из В на ось ON. Таким образом, мы получим точки F" и F", также лежащие на кривой (3.3.9).
Исходя из другой точки N, тем же самым образом получим новую четверку точек, лежащих на кривой (3.3.9). Исключение составит точка Ni = ?2/72- Исходя из нее, получим только две точки: К и L. Это будут нижняя и верхняя точки кривой (3.3.9).
Можно исходить не из значений N , а из значений N 2 . Направляясь от N 2 к кривой (3.3.11), поднимаясь затем до прямой У = сХ, а оттуда пересекая кривую (3.3.10), также найдем четыре точки кривой (3.3.9). Исключение составит точка No = ?1/71- Исходя из нее, получим только две точки: G и К. Это будут самая левая и самая правая точки кривой (3.3.9). Задавая разные N и N 2 и получив достаточно много точек, соединив их, приближенно построим кривую (3.3.9).
Из построения видно, что эго замкнутая кривая, содержащая внутри себя точку 12 = (?2/721 ?1/71)» исходящая из определенных начальных данных N ю и N20. Взяв другое значение С, т.е. другие начальные данные, получим другую замкнутую кривую, не пересекающую первую и также содержащую точку (?2/721 ?1/71)1 внутри себя. Таким образом, семейство траекторий (3.3.9) есть семейство замкнутых линий, окружающих точку 12 (см. рис. 3.3). Исследуем тип устойчивости этой особой точки, воспользовавшись методом Ляпунова.
Так как все параметры е 1, ?2, 71,72 положительны, точка (N[расположена в положительном квадранте фазовой плоскости. Линеаризация системы вблизи этой точки дает:

Здесь n(t) и 7i2(N1, N 2 :
Характеристическое уравнение системы (3.3.13):

Корни этого уравнения чисто мнимые:

Таким образом, исследование системы показывает, что траектории вблизи особой точки представлены концентрическими эллипсами, а сама особая точка - центр (рис. 3.4). Рассматриваемая модель Вольтерра и вдали от особой точки имеет замкнутые траектории, хотя форма этих траекторий уже отличается от эллипсоидальной. Поведение переменных Ni, N 2 во времени показано на рис. 3.5.

Рис. 3.4.

Рис. 3.5. Зависимость численности жертвы N i и хищника N 2 от времени
Особая точка типа центр устойчива, но не асимптотически. Покажем на данном примере, в чем это заключается. Пусть колебания Ni(t) и ЛГгМ происходят таким образом, что изображающая точка движется по фазовой плоскости по траектории 1 (см. рис. 3.4). В момент, когда точка находится в положении М, в систему извне добавляется некоторое количество особей N 2, такое, что изображающая точка переходит скачком из точки М в точку Л/". После этого, если система снова предоставлена самой себе, колебания Ni и N 2 уже будут происходить с большими амплитудами, чем прежде, и изображающая точка двигается по траектории 2. Это и означает, что колебания в системе неустойчивы: они навсегда изменяют свои характеристики при внешнем воздействии. В дальнейшем мы рассмотрим модели, описывающие устойчивые колебательные режимы, и покажем, что на фазовой плоскости такие асимптотические устойчивые периодические движения изображаются при помощи предельных циклов.
На рис. 3.6 изображены экспериментальные кривые - колебания численности пушных зверей в Канаде (по данным компании Гудзонова залива). Эти кривые построены на основании данных по числу заготовленных шкурок. Периоды колебаний численности зайцев (жертв) и рысей (хищников) примерно одинаковы и порядка 9 10 лет. При этом максимум численности зайцев опережает, как правило, максимум численности рысей на один год.
Форма этих экспериментальных кривых значительно менее правильная, чем теоретических. Однако в данном случае достаточно того, что модель обеспечивает совпадение наиболее существенных характеристик теоретических и экспериментальных кривых, г.е. величин амплитуды и сдвига фаз между колебаниями численностей хищников и жертв. Гораздо более серьезным недостатком модели Вольтерра является неустойчивость решений системы уравнений. Действительно, как уже говорилось выше, любое случайное изменение численности того или другого вида должно привести, следуя модели, к изменению амплитуды колебаний обоих видов. Естественно, что в природных условиях животные подвергаются бесчисленному количеству таких случайных воздействий. Как видно из экспериментальных кривых, амплитуда колебаний численностей видов мало изменяется от года к году.
Модель Вольтерра - эталонная (базовая) для математической экологии в той же мере, в какой модель гармонического осциллятора является базовой для классической и квантовой механики. При помощи этой модели на основе очень упрощенных представлений о характере закономерностей, описывающих поведение системы, сугубо математи-
Глава 3

Рис. 3.6. Кинетические кривые численности пушных зверей по данным пуш ной компании Гудзонова залива (Сетон-Томсон, 1987) ческими средствами было выведено заключение о качественном характере поведения такой системы - о наличии в такой системе колебаний численности популяции. Без построения математической модели и ее использования такой вывод был бы невозможен.
В рассмотренном нами выше самом простом виде системе Воль- терра присущи два принципиальных и взаимосвязанных недостатка. Их «устранению» посвящена обширная эколого-математическая литература. Во-первых, включение в модель любых, сколь угодно малых, дополнительных факторов качественным образом меняет поведение системы. Второй «биологический» недостаток модели заключается в том, что в нее не включены принципиальные свойства, присущие любой паре взаимодействующих по принципу хищник-жертва популяций: эффект насыщения хищника, ограниченность ресурсов хищника и жертвы даже при избытке жертвы, возможность минимальной численности жертв, доступных для хищника, и пр.
С целью устранения этих недостатков были предложены разными авторами различные модификации системы Вольтерра. Наиболее ин- тересные из них будут рассмотрены в разделе 3.5. Здесь остановимся лишь на модели, учитывающей самоограничения в росте обеих популяций. На примере этой модели наглядно видно, как может меняться характер решений при изменении параметров системы.
Итак, рассматривается система

Система (3.3.15) отличается от ранее рассмотренной системы (3.3.6) наличием в правых частях уравнений членов вида -7uNf,
Эти члены отражают тот факт, что численность популяции жертв не может расти до бесконечности даже в отсутствие хищников в силу ограниченности пищевых ресурсов, ограниченности ареала существования. Такие же «самоограничения» накладываются и на популяцию хищников.
Для нахождения стационарных численностей видов iVi и N 2 приравняем к нулю правые части уравнений системы (3.3.15). Решения с нулевыми значениями численностей хищников или жертв не будут нас сейчас интересовать. Поэтому рассмотрим систему алгебраических
уравнений
 Ее решение
Ее решение
дает нам координаты особой точки. На параметры системы здесь следует положить условие положительности стационарных численностей: N > 0 и N 2 > 0. Корни характеристического уравнения системы, линеаризованной в окрестности особой точки (3.3.16):

Из выражения для характеристических чисел видно, что если выполнено условие
то численности хищников и жертв совершают во времени затухающие колебания, система имеет ненулевую особую точку устойчивый фокус. Фазовый портрет такой системы изображен на рис. 3.7 а.
Допустим, что параметры в неравенстве (3.3.17) так изменяют свои значения, что условие (3.3.17) обращается в равенство. Тогда характеристические числа системы (3.3.15) равны, а ее особая точка будет лежать на границе между областями устойчивых фокусов и узлов. При изменении знака неравенства (3.3.17) на обратный особая точка становится устойчивым узлом. Фазовый портрет системы для этот случая представлен на рис. 3.76.
Как и в случае одной популяции, для модели (3.3.6) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения JVj, N 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке


Рис. 3.7. Фазовый портрет системы (3.3.15): а - при выполнении соотношения (3.3.17) между параметрами; б - при выполнении обратного соотношения между параметрами
фазовой траектории какая-либо переменная не очень велика, то существенное значение могут приобрести случайные флуктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида. Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический «дрейф» рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник- жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934; 2000), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum - хищника. Ожидавшиеся согласно детерминистическим уравнениям (3.3.6) равновесные численности в этих экспериментах составляли примерно всего но пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники).
Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. В модели хищник жертва незатухающие колебания появляются вследствие выбора специальной формы уравнений модели (3.3.6). При этом модель становится негрубой, что свидетельствует об отсутствии в такой системе механизмов, стремящихся сохранить ее состояние. Однако в природе и эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде. Рассмотрению таких обобщенных моделей посвящен раздел 3.5.
Приспособления, вырабатываемые жертвами для противодействия хищникам , способствуют выработке у хищников механизмов преодоления этих приспособлений. Длительное совместное существование хищников и жертв приводит к формированию системы взаимодействия, при которой обе группы устойчиво сохраняются на изучаемой территории. Нарушение такой системы часто приводит к отрицательным экологическим последствиям.
Негативное влияние нарушения коэволюционных связей наблюдается при интродукции видов. В частности, козы и кролики , интродуцированные в Австралии , не имеют на этом материке эффективных механизмов регуляции численности, что приводит к разрушению природных экосистем .
Математическая модель
Допустим, что на некоторой территории обитают два вида животных : кролики (питающиеся растениями) и лисы (питающиеся кроликами). Пусть число кроликов , число лис . Используя Модель Мальтуса с необходимыми поправками, учитывающими поедание кроликов лисами, приходим к следующей системе, носящей имя модели Вольтерры - Лотки :
\begin{cases} \dot x=(\alpha -c y)x;\\
\dot y=(-\beta+d x) y. \end{cases}
Поведение модели
Групповой образ жизни хищников и их жертв радикально меняет поведение модели, придает ей повышенную устойчивость.
Обоснование: при групповом образе жизни снижается частота случайных встреч хищников с потенциальными жертвами, что подтверждается наблюдениями за динамикой численности львов и антилоп гну в парке Серенгети .
История
Модель совместного существования двух биологических видов (популяций) типа «хищник - жертва» называется также моделью Вольтерры - Лотки.
См. также
Напишите отзыв о статье "Система «хищник - жертва»"
Примечания
Литература
- В. Вольтерра, Математическая теория борьбы за существование. Пер. с франц. О. Н. Бондаренко. Под ред и послесловием Ю. М. Свирежева. М.: Наука, 1976. 287 c. ISBN 5-93972-312-8
- А. Д. Базыкин, Математическая биофизика взаимодействующих популяций. М.: Наука, 1985. 181 с.
- А. Д. Базыкин, Ю. А. Кузнецов, А. И. Хибник, Портреты бифуркаций (Бифуркационные диаграммы- динамических систем на плоскости) /Серия «Новое в жизни, науке, технике. Математика, кибернетика» - М.: Знание, 1989. 48 с.
- П. В. Турчин,
Ссылки
Отрывок, характеризующий Система «хищник - жертва»
– Charmant, charmant, [Прелестно, прелестно,] – сказал князь Василий.– C"est la route de Varsovie peut etre, [Это варшавская дорога, может быть.] – громко и неожиданно сказал князь Ипполит. Все оглянулись на него, не понимая того, что он хотел сказать этим. Князь Ипполит тоже с веселым удивлением оглядывался вокруг себя. Он так же, как и другие, не понимал того, что значили сказанные им слова. Он во время своей дипломатической карьеры не раз замечал, что таким образом сказанные вдруг слова оказывались очень остроумны, и он на всякий случай сказал эти слова, первые пришедшие ему на язык. «Может, выйдет очень хорошо, – думал он, – а ежели не выйдет, они там сумеют это устроить». Действительно, в то время как воцарилось неловкое молчание, вошло то недостаточно патриотическое лицо, которого ждала для обращения Анна Павловна, и она, улыбаясь и погрозив пальцем Ипполиту, пригласила князя Василия к столу, и, поднося ему две свечи и рукопись, попросила его начать. Все замолкло.
– Всемилостивейший государь император! – строго провозгласил князь Василий и оглянул публику, как будто спрашивая, не имеет ли кто сказать что нибудь против этого. Но никто ничего не сказал. – «Первопрестольный град Москва, Новый Иерусалим, приемлет Христа своего, – вдруг ударил он на слове своего, – яко мать во объятия усердных сынов своих, и сквозь возникающую мглу, провидя блистательную славу твоея державы, поет в восторге: «Осанна, благословен грядый!» – Князь Василий плачущим голосом произнес эти последние слова.
Билибин рассматривал внимательно свои ногти, и многие, видимо, робели, как бы спрашивая, в чем же они виноваты? Анна Павловна шепотом повторяла уже вперед, как старушка молитву причастия: «Пусть дерзкий и наглый Голиаф…» – прошептала она.
Князь Василий продолжал:
– «Пусть дерзкий и наглый Голиаф от пределов Франции обносит на краях России смертоносные ужасы; кроткая вера, сия праща российского Давида, сразит внезапно главу кровожаждущей его гордыни. Се образ преподобного Сергия, древнего ревнителя о благе нашего отечества, приносится вашему императорскому величеству. Болезную, что слабеющие мои силы препятствуют мне насладиться любезнейшим вашим лицезрением. Теплые воссылаю к небесам молитвы, да всесильный возвеличит род правых и исполнит во благих желания вашего величества».
– Quelle force! Quel style! [Какая сила! Какой слог!] – послышались похвалы чтецу и сочинителю. Воодушевленные этой речью, гости Анны Павловны долго еще говорили о положении отечества и делали различные предположения об исходе сражения, которое на днях должно было быть дано.
– Vous verrez, [Вы увидите.] – сказала Анна Павловна, – что завтра, в день рождения государя, мы получим известие. У меня есть хорошее предчувствие.
Предчувствие Анны Павловны действительно оправдалось. На другой день, во время молебствия во дворце по случаю дня рождения государя, князь Волконский был вызван из церкви и получил конверт от князя Кутузова. Это было донесение Кутузова, писанное в день сражения из Татариновой. Кутузов писал, что русские не отступили ни на шаг, что французы потеряли гораздо более нашего, что он доносит второпях с поля сражения, не успев еще собрать последних сведений. Стало быть, это была победа. И тотчас же, не выходя из храма, была воздана творцу благодарность за его помощь и за победу.
Предчувствие Анны Павловны оправдалось, и в городе все утро царствовало радостно праздничное настроение духа. Все признавали победу совершенною, и некоторые уже говорили о пленении самого Наполеона, о низложении его и избрании новой главы для Франции.
Вдали от дела и среди условий придворной жизни весьма трудно, чтобы события отражались во всей их полноте и силе. Невольно события общие группируются около одного какого нибудь частного случая. Так теперь главная радость придворных заключалась столько же в том, что мы победили, сколько и в том, что известие об этой победе пришлось именно в день рождения государя. Это было как удавшийся сюрприз. В известии Кутузова сказано было тоже о потерях русских, и в числе их названы Тучков, Багратион, Кутайсов. Тоже и печальная сторона события невольно в здешнем, петербургском мире сгруппировалась около одного события – смерти Кутайсова. Его все знали, государь любил его, он был молод и интересен. В этот день все встречались с словами:
– Как удивительно случилось. В самый молебен. А какая потеря Кутайсов! Ах, как жаль!
– Что я вам говорил про Кутузова? – говорил теперь князь Василий с гордостью пророка. – Я говорил всегда, что он один способен победить Наполеона.
Но на другой день не получалось известия из армии, и общий голос стал тревожен. Придворные страдали за страдания неизвестности, в которой находился государь.
– Каково положение государя! – говорили придворные и уже не превозносили, как третьего дня, а теперь осуждали Кутузова, бывшего причиной беспокойства государя. Князь Василий в этот день уже не хвастался более своим protege Кутузовым, а хранил молчание, когда речь заходила о главнокомандующем. Кроме того, к вечеру этого дня как будто все соединилось для того, чтобы повергнуть в тревогу и беспокойство петербургских жителей: присоединилась еще одна страшная новость. Графиня Елена Безухова скоропостижно умерла от этой страшной болезни, которую так приятно было выговаривать. Официально в больших обществах все говорили, что графиня Безухова умерла от страшного припадка angine pectorale [грудной ангины], но в интимных кружках рассказывали подробности о том, как le medecin intime de la Reine d"Espagne [лейб медик королевы испанской] предписал Элен небольшие дозы какого то лекарства для произведения известного действия; но как Элен, мучимая тем, что старый граф подозревал ее, и тем, что муж, которому она писала (этот несчастный развратный Пьер), не отвечал ей, вдруг приняла огромную дозу выписанного ей лекарства и умерла в мучениях, прежде чем могли подать помощь. Рассказывали, что князь Василий и старый граф взялись было за итальянца; но итальянец показал такие записки от несчастной покойницы, что его тотчас же отпустили.
система РА88, которая одновременно предсказывает вероятность более чем 100 фармакологических эффектов и механизмов действия вещества на основе его структурной формулы. Эффективность применения этого подхода к планированию скрининга составляет около 800%, а точность компьютерного прогноза на 300% превосходит предсказание экспертов.
Итак, одним из конструктивных инструментов получения новых знаний и решений в медицине является метод математического моделирования. Процесс математизации медицины – частое проявление взаимопроникновения научных знаний, повышающее эффективность лечебно-профилактической работы.
4. Математическая модель «хищники-жертвы»
Впервые в биологии математическую модель периодического изменения числа антагонистических видов животных предложил итальянский математик В. Вольтерра с сотрудниками. Модель, предложенная Вольтерра, явилась развитием идеи, намеченной в 1924 году А. Лоттки в книге "Элементы физической биологии". Поэтому эта классическая математическая модель известна как модель "Лоттки-Вольтерра".
Хотя в природе отношения антагонистических видов более сложные, чем в модели, тем не менее они являются хорошей учебной моделью, на которой можно изучать основные идеи математического моделирования.
Итак, задача : в некотором экологически замкнутом районе живут два вида животных (например, рыси и зайцы). Зайцы (жертвы) питаются растительной пищей, имеющейся всегда в достаточном количестве (в рамках данной модели не учитывается ограниченность ресурсов растительной пищи). Рыси (хищники) могут питаться только зайцами. Необходимо определить, как будет меняться численность жертв и хищников с течением времени в такой экологической системе. Если популяция жертв увеличивается, вероятность встреч хищников с жертвами возрастает, и, соответственно, после некоторой временной задержки, растет популяция хищников. Эта достаточно простая модель вполне адекватно описывает взаимодействие между реальными популяциями хищников и жертв в природе.
Теперь приступим к составлению дифференциальных уравнений. Обо-
значим число жертв через N, а число хищников через M. Числа N и M являются функциями времени t . В нашей модели учтем следующие факторы:
а) естественное размножение жертв; б) естественная гибель жертв;
в) уничтожение жертв за счет поедания их хищниками; г) естественное вымирание хищников;
д) увеличение числа хищников за счет размножения при наличии пищи.
Так как речь идет о математической модели, то задачей является получение уравнений, в которые входили бы все намеченные факторы и которые описывали бы динамику, то есть изменение числа хищников и жертв со временем.
Пусть за некоторое время t количество жертв и хищников изменится на ∆N и ∆M. Изменение числа жертв ∆N за время ∆t определяется, во-первых, увеличением в результате естественного размножения (которое пропорционально имеющемуся количеству жертв):
где В – коэффициент пропорциональности, характеризующий скорость естественного вымирания жертв.
В основе вывода уравнения, описывающего уменьшение числа жертв изза поедания их хищниками, лежит идея о том, что чем чаще происходит их встреча, тем быстрее уменьшается число жертв. Ясно также, что частота встреч хищников с жертвой пропорционально и числу жертв и числу хищников, то
Поделив левую и правую часть уравнения (4) на ∆t и перейдя к пределу при ∆t→0 , получим дифференциальное уравнение первого порядка:
Для того, чтобы решить это уравнение, нужно знать, как меняется число хищников (М ) со временем. Изменение числа хищников (∆М ) определяется увеличением из-за естественного размножения при наличии достаточного количества пищи (М 1 = Q∙N∙M∙∆t ) и уменьшением из-за естественного вымирания хищников (M 2 = - P∙M∙∆t ):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Из уравнения (6) можно получить дифференциальное уравнение:
Дифференциальные уравнения (5) и (7) представляют собой математическую модель "хищники-жертвы". Достаточно определить значения коэффици-

ентов A, B, C, Q, P и математическую модель можно использовать для решения поставленной задачи.
Проверка и корректировка математической модели. В данной лабора-
торной работе предлагается кроме просчета наиболее полной математической модели (уравнения 5 и 7), исследовать более простые, в которых что-либо не учитывается.
Рассмотрев пять уровней сложности математической модели, можно "почувствовать" этап проверки и корректировки модели.
1-ый уровень – в модели учтено для "жертв" только их естественное размножение, "хищники" отсутствуют;
2-ой уровень – в модели учтено для "жертв" их естественное вымирание, "хищники" отсутствуют;
3-ий уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, "хищники" отсутствуют;
4-ый уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, а также поедание "хищниками", но число "хищников" остается неизменным;
5-ый уровень – в модели учтены все обсуждаемые факторы.
Итак, имеем следующую систему дифференциальных уравнений:
где М – число "хищников"; N – число "жертв";
t – текущее время;
A – скорость размножения "жертв"; C – частота встреч "хищники-жертвы"; B – скорость вымирания "жертв";
Q – размножение "хищников";
P – вымирание "хищников".
1-ый уровень: М = 0, В = 0; 2-ой уровень: М = 0, А = 0; 3-ий уровень: М = 0; 4-ый уровень: Q = 0, Р = 0;
5-ый уровень: полная система уравнений.
Подставляя значения коэффициентов в каждый уровень, будем получать разные решения, например:
Для 3-его уровня значение коэффициента М=0 , тогда

решая уравнение получим
Аналогично для 1-го и 2-го уровней. Что касается 4-го и 5-го уровней, то здесь необходимо решать систему уравнений методом Рунге-Кутта. В результате получим решение математических моделей данных уровней.
II. РАБОТА СТУДЕНТОВ ВО ВРЕМЯ ПРАКТИЧЕСКОГО ЗАНЯТИЯ
Задание 1 . Устно-речевой контроль и коррекция усвоения теоретического материала занятия. Сдача допуска к занятию.
Задание 2 . Выполнение лабораторной работы, обсуждение полученных результатов, оформление конспекта.
Выполнение работы
1. С рабочего стола компьютера вызвать программу "Лаб. №6", щелкнув по соответствующему ярлыку два раза левой клавишей "мыши".
2. Щелкнуть дважды левой клавишей "мыши" по ярлыку "PREDATOR".
3. Выбрать ярлык "PRED" и повторить вызов программы левой клавишей "мыши" (щелкнув дважды).
4. После титульной заставки нажать "ENTER".
5. Моделирование начинать с 1-го уровня.
6. Ввести год, с которого будет проводиться анализ модели: например, 2000
7. Выбрать временные интервалы, например, в течение 40 лет, через 1 год (затем через 4 года).
2-ой уровень: B = 0.05; N0 = 200;
3-ий уровень: A = 0.02; B = 0.05; N = 200;
4-ый уровень: A = 0.01; B = 0.002; C = 0.01; N0 = 200; M = 40; 5-ый уровень: A = 1; B = 0.5; C = 0.02; Q = 0.002; P = 0.3; N0 = 200;
9. Подготовить письменный отчет по работе, который должен содержать уравнения, графики, результаты расчета характеристик модели, выводы по проделанный работе.
Задание 3. Контроль конечного уровня знаний:
а) устно-речевой отчет за выполненную лабораторную работу; б) решение ситуационных задач; в) компьютерное тестирование.
Задание 4. Задание на следующее занятие: раздел и тема занятия, согласование тем реферативных докладов (объем доклада 2-3 стр., регламент 5-7 мин.).
Модели взаимодействия двух видов
Гипотезы Вольтерра. Аналогии с химической кинетикой. Вольтерровские модели взаимодействий. Классификация типов взаимодействий Конкуренция. Хищник-жертва. Обобщенные модели взаимодействия видов. Модель Колмогорова. Модель взаимодействия двух видов насекомых Макартура. Параметрический и фазовые портреты системы Базыкина.
Основателемсовременной математической теории популяций справедливо считается итальянский математик Вито Вольтерра, разработавший математическую теорию биологических сообществ, аппаратом которой служат дифференциальные и интегро-дифференциальные уравнения. (Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris , 1931). В последующие десятилетия популяционная динамика развивалась, в основном, в русле высказанных в этой книге идей. Русский перевод книги Вольтерра вышел в 1976 г. под названием: «Математическая теория борьбы за существование» с послесловием Ю.М. Свирежева, в котором рассматривается история развития математической экологии в период 1931‑1976 гг.
Книга Вольтерра написана так, как пишут книги по математике. В ней сначала сформулированы некоторые предположения о математических объектах, которые предполагается изучать, а затем проводится математическое исследование свойств этих объектов.
Системы, изученные Вольтерра, состоят их двух или нескольких видов. В отдельных случаях рассматривается запас используемой пищи. В основу уравнений, описывающих взаимодействие этих видов, положены следующие представления.
Гипотезы Вольтерра
1. Пища либо имеется в неограниченном количестве, либо ее поступление с течением времени жестко регламентировано.
2. Особи каждого вида отмирают так, что в единицу времени погибает постоянная доля существующих особей.
3. Хищные виды поедают жертв, причем в единицу времени количество съеденных жертв всегда пропорционально вероятности встречи особей этих двух видов, т.е. произведению количества хищников на количество жертв.
4. Если имеется пища в ограниченном количестве и несколько видов, которые способны ее потреблять, то доля пищи, потребляемой видом в единицу времени, пропорциональна количеству особей этого вида, взятому с некоторым коэффициентом, зависящим от вида (модели межвидовой конкуренции).
5. Если вид питается пищей, имеющейся в неограниченном количестве, прирост численности вида в единицу времени пропорционален численности вида.
6. Если вид питается пищей, имеющейся в ограниченном количестве, то его размножение регулируется скоростью потребления пищи, т.е. за единицу времени прирост пропорционален количеству съеденной пищи.
Аналогии с химической кинетикой
Эти гипотезы имеют близкие параллели с химической кинетикой. В уравнениях популяционной динамики, как и в уравнениях химической кинетики, используется “принцип соударений”, когда скорость реакции пропорциональна произведению концентраций реагирующих компонентов.
Действительно, согласно гипотезам Вольтерра, скорость процесса отмирания каждого вида пропорциональна численности вида. В химической кинетике это соответствует мономолекулярной реакции распада некоторого вещества, а в математической модели – отрицательным линейным членам в правых частях уравнений.
Согласно представлениям химической кинетики, скорость бимолекулярной реакции взаимодействия двух веществ пропорциональна вероятности столкновения этих веществ, т.е. произведению их концентрации. Точно так же, в соответствии с гипотезами Вольтерра, скорость размножения хищников (гибели жертв) пропорциональна вероятности встреч особей хищника и жертвы, т.е. произведению их численностей. И в том и в другом случае в модельной системе появляются билинейные члены в правых частях соответствующих уравнений.
Наконец, линейные положительные члены в правых частях уравнений Вольтерра, отвечающие росту популяций в неограниченных условиях, соответствуют автокаталитическим членам химических реакций. Такое сходство уравнений в химических и экологических моделях позволяет применить для математического моделирования кинетики популяций те же методы исследований, что и для систем химических реакций.
Классификация типов взаимодействий
В соответствии с гипотезами Вольтерра взаимодействие двух видов, численности которых x 1 иx 2 , могут быть описаны уравнениями:
(9.1)
Здесь параметры a i ‑ константы собственной скорости роста видов, c i ‑ константы самоограничения численности (внутривидовой конкуренции), b ij ‑ константы взаимодействия видов, (i , j= 1,2). Знаки этих коэффициентов определяют тип взаимодействия.
В биологической литературе обычно классифицируют взаимодействия по участвующим в них механизмам. Разнообразие здесь огромно: различные трофические взаимодействия, химические взаимодействия, существующие между бактериями и планктонными водорослями, взаимодействия грибов с другими организмами, сукцессии растительных организмов, связанные в частности, с конкуренцией за солнечный свет и с эволюцией почв и т.д. Такая классификация кажется необозримой.
E . Одум, учитывая предложенные В.Вольтерра модели, предложил классификацию не по механизмам, а по результатам. Согласно этой классификации, оценивать взаимоотношения следует как положительные, отрицательные или нейтральные в зависимости от того, возрастает, убывает или остается неизменной численность одного вида в присутствии другого вида. Тогда основные типы взаимодействий могут быть представлены в виде таблицы.
ТИПЫ ВЗАИМОДЕЙСТВИЯ ВИДОВ
|
СИМБИОЗ |
b 12 ,b 21 >0 |
||
|
КОММЕНСАЛИЗМ |
b 12 ,>0, b 21 =0 |
||
|
ХИЩНИК-ЖЕРТВА |
b 12 ,>0, b 21 <0 |
||
|
АМЕНСАЛИЗМ |
b 12 ,=0, b 21 <0 |
||
|
КОНКУРЕНЦИЯ |
b 12 , b 21 <0 |
||
|
НЕЙТРАЛИЗМ |
b 12 , b 21 =0 |
В последнем столбце указаны знаки коэффициентов взаимодействия из системы (9.1)
Рассмотрим основные типы взаимодействий
УравненияКОНКУРЕНЦИИ:
Как мы уже видели в лекции 6, уравнения конкуренции имеют вид:
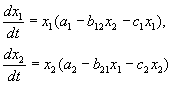 (9.2)
(9.2)
Стационарные решения системы:
(1).
![]()
Начало координат, при любых параметрах системы представляет собой неустойчивый узел.
(2).
![]() (9.3)
(9.3)
C тационарное состояние (9.3) представляет собой седло при a 1 >b 12 /с 2 и
устойчивый узел приa 1 12 /с 2 . Это условие означает, что вид вымирает, если его собственная скорость роста меньше некоторой критической величины.
(3).
![]() (9.4)
(9.4)
C тационарное решение (9.4) ¾ седло при a 2 >b 21 /c 1 и устойчивый узел при a 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Стационарное состояние (9.5) характеризует сосуществование двух конкурирующих видов и представляет собой устойчивый узел в случае выполнения соотношения:
![]()
Отсюда следует неравенство:
b
12
b
21
позволяющее сформулировать условие сосуществования видов:
Произведение коэффициентов межпопуляционного взаимодействия меньше произведения коэффициентов внутри популяционного взаимодействия.
Действительно, пусть естественные скорости роста двух рассматриваемых видов a 1 , a 2 одинаковы. Тогда необходимым для устойчивости условием будет
c 2 > b 12 ,c 1 > b 21 .
Эти неравенства показывают, что увеличение численности одного из конкурентов сильнее подавляет его собственный рост, чем рост другого конкурента. Если численность обоих видов ограничивается, частично или полностью, различными ресурсами, приведенные выше неравенства справедливы. Если же оба вида имеют совершенно одинаковые потребности, то один из них окажется более жизнеспособным и вытеснит своего конкурента.
Поведение фазовых траекторий системы дает наглядное представление о возможных исходах конкуренции. Приравняем нулю правые части уравнений системы (9.2):
x 1 (a 1 –c 1 x 1 – b 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (a 2 –b 21 x 1 – c 2 x 2) = 0 (dx 2 /dt = 0),
При этом получим уравнения для главных изоклин системы
x 2 = – b 21 x 1 / c 2 +a 2 /c 2 , x 2 = 0
– уравнения изоклин вертикальных касательных.
x 2 = – c 1 x 1 / b 12 +a 1 /b 12 , x 1 = 0
– уравнения изоклин вертикальных касательных.
Точки попарного пересечения изоклин вертикальных и горизонтальных касательных
систем представляют собой стационарные решения системы уравнений (9.2.), а их
координаты ![]() суть стационарные численности конкурирующих видов.
суть стационарные численности конкурирующих видов.
Возможное расположение главных изоклин в системе (9.2) изображено на рис.9.1. Рис. 9.1 а соответствует выживанию вида x 1 , рис. 9.1 б – выживанию вида x 2 , рис. 9.1 в – сосуществованию видов при выполнении условия (9.6). Рисунок 9.1 г демонстрирует триггерную систему. Здесь исход конкуренции зависит от начальных условий. Ненулевое для обоих видов стационарное состояние (9.5) – неустойчивое. Это – седло, через которое проходит сепаратриса, отделяющая области выживания каждого из видов.

Рис. 9.1. Расположение главных изоклин на фазовом портрете вольтерровской системы конкуренции двух видов (9.2) при разном соотношении параметров. Пояснения в тексте.
Для изучения конкуренции видов ставились эксперименты на самых различных организмах. Обычно выбирают два близкородственных вида и выращивают их вместе и по отдельности в строго контролируемых условиях. Через определенные промежутки времени проводят полный или выборочный учет численности популяции. Регистрируют данные по нескольким повторным экспериментам и анализируют. Исследования проводили на простейших (в частности, инфузориях), многих видах жуков рода Tribolium, дрозофиллах, пресноводных ракообразных (дафниях). Много экспериментов проводилось на микробных популяциях (см. лекция 11). В природе также проводили эксперименты, в том числена планариях (Рейнольдс), двух видах муравьев (Понтин) и др. На рис. 9.2. изображены кривые роста диатомовых водорослей, использующих один и тот же ресурс (занимающих одну и ту же экологическую нишу). При выращивании в монокультуре Asterionella Formosa выходит на постоянный уровень плотности и поддержвает концентрацию ресурса (силиката) на постоянно низком уровне. Б. При выращивании в монокультуре Synedrauina ведет себя сходным образом и поддерживает концентрацию силиката на еще более низком уровне. В. При совместном культивировании (в двух повторностях) Synedrauina вытесняет Asterionella Formosa . По-видимому, Synedra

Рис. 9.2. Конкуренция у диатомовых водорослей. а ‑ при выращивании в монокультуре Asterionella Formosa выходит на постоянный уровень плотности и поддерживает концентрацию ресурса (силиката) на постоянно низком уровне.б ‑ при выращивании в монокультуре Synedrauina ведет себя сходным образом и поддерживает концентрацию силиката на еще более низком уровне. в ‑ при совместном культивировании (в двух повторностях) Synedruina вытесняет Asterionella Formosa . По-видимому, Synedra выигрывает конкуренцию благодаря своей способности к более полному использованию субстрата (см. также Лекцию 11).
Широко известны эксперименты по изучению конкуренции Г. Гаузе, продемонстрировавшие выживание одного из конкурирующих видов и позволившие ему сформулировать «закон конкурентного исключения». Закон гласит, что в одной экологической нише может существовать только один вид. На рис. 9.3. приведены результаты экспериментов Гаузе для двух видов Parametium , занимающих одну экологическую нишу (рис. 9.3 а, б) и видами, занимающими разные экологические ниши (рис. 9.3. в).

Рис. 9.3. а - Кривые роста популяций двух видов Parametium в одновидовыхкультурах. Черные кружки – P Aurelia , белые кружки – P . Caudatum
б - Кривые роста P Aurelia и P . Caudatum в смешанной культуре.
По Gause , 1934
Модель конкуренции (9.2) имеет недостатки, в частности, из нее следует, что сосуществование двух видов возможно лишь в случае, если их численность ограничивается разными факторами, но модель не дает указаний, насколько велики должны быть различия для обеспечения длительного сосуществования. В то же время известно, что для длительного сосуществования в изменчивой среде необходимо различие, достигающее определенной величины. Внесение в модель стохастических элементов (например, введение функции использования ресурса) позволяет количественно исследовать эти вопросы.
Система ХИЩНИК+ЖЕРТВА
 (9.7)
(9.7)
Здесь, в отличие от (9.2) знаки b 12 и b 21 - разные. Как и в случае конкуренции, начало координат
![]() (9.8)
(9.8)
является особой точкой типа неустойчивый узел. Три других возможных стационарных состояния:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Таким образом, возможно выживание только жертвы (9.10), только хищника (9.9) (если у него имеются и другие источники питания) и сосуществование обоих видов (9.11). Последний вариант уже был рассмотрен нами в лекции 5. Возможные типы фазовых портретов для системы хищник-жертва представлены на рис. 9.4.
Изоклины горизонтальных касательных представляют собой прямые
x 2 = – b 21 х 1 /c 2 + a 1 /c 2 , х 2 = 0,
а изоклины вертикальных касательных – прямые
x 2 = – c 1 х 1 /b 12 + a 2 /b 12 , х 1 = 0.
Стационарные точки лежат на пересечении изоклин вертикальных и горизонтальных касательных.

Из рис. 9.4 видно следующее. Система хищник – жертва (9.7) может иметь устойчивое положение равновесия, в кот o ром популяция жертв полностью вымерла ( ) и остались только хищники (точка 2 на рис. 9.4 а). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х 1 хищник х 2 – имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для х 2 . Особые точки (1) и (3) (рис. 9.4 а ) являются неустойчивыми. Вторая возможность – устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы – устойчивая точка (3) (рис. 9.4 6 ). Здесь особая точка (1) – также неустойчивый узел.
Наконец, третья возможность – устойчивое сосуществование популяций хищника и жертвы (рис. 9.4 в ), стационарные численности которых выражаются формулами (9.11).
Как и в случае одной популяции (см. Лекция 3), для модели (9.7) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения x 1 , x 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке фазовой траектории какая–либо переменная не очень велика, то существенное значение могут приобрести случайные флюктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида.
Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический “дрейф” рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник – жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum – хищника. Ожидавшиеся согласно детерминистическим уравнениям (9.7) равновесные численности в этих экспериментах составляли примерно всего по пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники) Результаты экспериментов представлены на рис. 9.5.

Рис . 9.5. Рост Parametium caudatum и хищной инфузории Dadinium nasutum . Из : Gause G.F. The struggle for existence. Baltimore , 1934
Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. Однако в природе и в эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде.
Обобщенные модели взаимодействия двух видов
Было предложено большое число моделей, описывающих взаимодействие видов, правые части уравнений которых представляли собой функции численностей взаимодействующих популяций. Решался вопрос о выработке общих критериев, позволяющих установить, какого вида функции могут описать особенности поведения временного численности популяции, в том числе устойчивые колебания. Наиболее известные из этих моделей принадлежат Колмогорову (1935, переработанная статья ‑ 1972) и Розенцвейгу (1963).
 (9.12)
(9.12)
В модель заложены следующие предположения:
1) Хищники не взаимодействуют друг с другом, т.е. коэффициент размножения хищников k 2 и число жертв L , истребляемых в единицу времени одним хищником, не зависит от y.
2) Прирост числа жертв при наличии хищников равен приросту в отсутствие хищников минус число жертв, истребляемых хищниками. Функции k 1 (x ), k 2 (x ), L (x ), ‑ непрерывны и определены на положительной полуоси x , y ³ 0.
3) dk 1 /dx< 0. Это означает, что коэффициент размножения жертв в отсутствие хищника монотонно убывает с возрастанием численности жертв, что отражает ограниченность пищевых и иных ресурсов.
4) dk 2 / dx> 0, k 2 (0) < 0 < k 2 (¥ ). С ростом численности жертв коэффициент размножения хищников монотонно убывает с возрастанием численности жертв, переходя от отрицательных значений, (когда нечего есть) к положительным.
5) Число жертв, истребляемых одним хищником в единицу времени L (x )> 0 при N> 0; L (0)=0.
Возможные типы фазовых портретов системы (9.12) представлены на рис. 9.6:

Рис. 9.6. Фазовые портреты системы Колмогорова (9.12), описывающей взаимодействие двух видов при разных соотношениях параметров. Пояснения в тексте.
Стационарные решения (их два или три) имеют следующие координаты:
(1). ` x=0; ` y=0 .
Начало координат при любых значениях параметров представляет собой седло(рис. 9.6 а-г).
(2). ` x=A, ` y=0. (9.13)
A определяется из уравнения:
k 1 (A )=0.
Стационарное решение (9.13) ‑ седло, если B< A (рис. 9.6 а , б , г ), B определяется из уравнения
k 2 (B )=0
Точка (9.13) помещается в положительном квадранте, еслиB>A. Это ‑ устойчивый узел.
Последний случай, который соответствует гибели хищника и выживанию жертвы,представлен на рис. 9.6 в .
(3). ` x=B, ` y=C. (9.14)
Величина С определяется из уравнений:
Точка (9.14) – фокус (рис.9.6 а ) или узел (рис.9.6 г ), устойчивость которых зависит от знакавеличины s
s 2 = – k 1 (B) – k 1 (B )B+L (B )C.
Если s >0, точка устойчива, если s <0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 б )
В зарубежной литературе чаще рассматривается сходная модель, предложенная Розенцвейгом и Макартуром (1963):
 (9.15)
(9.15)
где f (x ) - скорость изменения численности жертв x в отсутствие хищников, Ф(x,y ) ‑ интенсивность хищничества, k ‑ коэффициент, характеризующий эффективность переработки биомассы жертвы в биомассу хищника, e - смертность хищника.
Модель (9.15) сводится к частному случаю модели Колмогорова (9.12) при следующих предположениях:
1) численность хищника ограничивается только численностью жертвы,
2) скорость, с которой данная особь хищника поедает жертву, зависит только от плотности популяции жертв и не зависит от плотности популяции хищников.
Тогда уравнения (9.15) принимают вид.

При описании взаимодействия реальных видов, правые части уравнений конкретизируются в соответствии с представлениями о биологических реалиях. Рассмотрим одну из самых популярных моделей такого типа.
Модельвзаимодействиядвух видов насекомых (M acArthur, 1971)
Модель, которую мы рассмотрим ниже, использовалась для решения практической задачи борьбы с вредными насекомыми с помощью стерилизации самцов одного из видов. Исходя из биологических особенностей взаимодействия видов, была написана следующая модель
 (9.16)
(9.16)
Здесь x,y - биомассы двух видов насекомых. Трофические взаимодействия описываемых в этой модели видов – весьма сложные. Этим определяется вид многочленов в правых частях уравнений.
Рассмотрим правую часть первого уравнения. Насекомые вида х поедают личинок вида у (член + k 3 y), но взрослые особи вида у поедают личинок вида х при условии высокой численности видов х или у или обоих видов (члены – k 4 xy, – у 2 ). При малых х смертность вида х выше, чем его естественный прирост (1–k 1 +k 2 x–x 2 < 0 при малых х). Во втором уравнении член k 5 отражает естественный прирост вида y; –k 6 y – самоограничение этого вида, –k 7 x – поедание личинок вида у насекомыми вида х, k 8 xy – прирост биомассы вида у за счет поедания взрослыми насекомыми вида у личинок вида х.
На рис. 9.7 представлен предельный цикл, являющийся траекторией устойчивого периодического решения системы (9.16).

Решение вопроса о том, каким образом обеспечить сосуществование популяции с ее биологическим окружением, разумеется, не может быть получено без учета специфики конкретной биологической системы и анализа всех ее взаимосвязей. Вместе с тем изучение формальных математических моделей позволяет ответить на некоторые общие вопросы. Можно утверждать, что для моделей типа (9.12) факт совместимости или несовместимости популяций не зависит от их начальной численности, а определяется только характером взаимодействия видов. Модель помогает ответить на вопрос: как следует воздействовать на биоценоз, управлять им, чтобы по возможности быстро уничтожить вредный вид.
Управление может сводиться к кратковременному, скачкообразному изменению величин численности х и у. Такой способ отвечает методам борьбы типа однократного уничтожения одной или обеих популяций химическими средствами. Из сформулированного выше утверждения видно, что для совместимых популяций этот метод борьбы будет малоэффективным, поскольку с течением времени система опять выйдет на стационарный режим.
Другой способ ‑ изменение вида функций взаимодействия между видами, например, при изменении значений параметров системы. Именно такому, параметрическому, способу отвечают биологические методы борьбы. Так при внедрении стерилизованных самцов уменьшается коэффициент естественного прироста популяции. Если при этом мы получим другой тип фазового портрета, такой, где имеется лишь устойчивое стационарное состояние с нулевой численностью вредителя, управление приведет к желаемому результату – уничтожению популяции вредного вида. Интересно отметить, что иногда воздействие целесообразно применить не к самому вредителю, а к его партнеру. Какой из способов более эффективен, в общем случае сказать нельзя. Это зависит от имеющихся в распоряжении средств управления и от явного вида функций, описывающих взаимодействие популяций.
Модель А.Д.Базыкина
Теоретический анализ моделей взаимодействий видов наиболее исчерпывающе проведен в книге А.Д.Базыкина “Биофизика взаимодействующих популяций” (М., Наука, 1985).
Рассмотрим одну из изученных в этой книге моделей типа хищник-жертва.
 (9.17)
(9.17)
Система (9.17) является обобщением простейшей модели хищник-жертва Вольтерра (5.17) с учетом эффекта насыщения хищников. В модели (5.17) предполагается, что интенсивность выедания жертв линейно растет с ростом плотности жертв, что при больших плотностях жертв не соответствует реальности. Для описания зависимости рациона хищника от плотности жертв могут быть выбраны разные функции. Наиболее существенно, чтобы выбранная функция с ростом x асимптотически стремилась к постоянному значению. В модели (9.6) использовалась логистическая зависимость. В модели Базыкина в роли такой функции выбрана гипербола x /(1+px ). Вспомним, что такой вид имеет формула Моно, описывающая зависимость скорость роста микроорганизмов от концентрации субстрата. Здесь в роли субстрата выступает жертва, а в роли микроорганизмов – хищник.
Система (9.17) зависит от семи параметров. Число параметров можно уменьшить с помощью замены переменных:
x ® (A/D )x; y ® (A/D )/y;
t ® (1/A )t; g (9.18)
и зависит от четырех параметров.
Для полного качественного исследования необходимо разбить четырехмерное пространство параметров на области с различным типом динамического поведения, т.е. построить параметрический, или структурный портрет системы.
Затем надо построить фазовые портреты для каждой из областей параметрического портрета и описать бифуркации, происходящие с фазовыми портретами на границах различных областей параметрического портрета.
Построение полного параметрического портрета производится в виде набора “срезов” (проекций) параметрического портрета малой размерности при фиксированных значениях некоторых из параметров.
Параметрический портрет системы (9.18) при фиксированных g и малых e представлен на рис.9.8. Портрет содержит 10 областей с различным типом поведения фазовых траекторий.
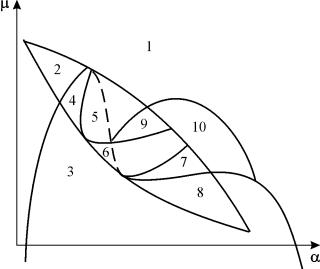
Рис. 9.8. Параметрический портрет системы (9.18) при фиксированных g
и малых e
Поведение системы при различных соотношениях параметров может быть существенно различным (рис.9.9). В системе возможны:
1) одно устойчивое равновесие (области 1 и 5);
2) один устойчивый предельный цикл (области 3 и 8);
3) два устойчивых равновесия (область 2)
4) устойчивый предельный цикл и неустойчивое равновесие внутри него (области 6, 7, 9, 10)
5) устойчивый предельный цикл и устойчивое равновесие вне его (область 4).
В параметрических областях 7, 9, 10 область притяжения равновесия ограничивается неустойчивым предельным циклом, лежащим внутри устойчивого. Наиболее интересно устроен фазовый портрет, соответствующий области 6 на параметрическом портрете. Детально он изображен на рис. 9.10.
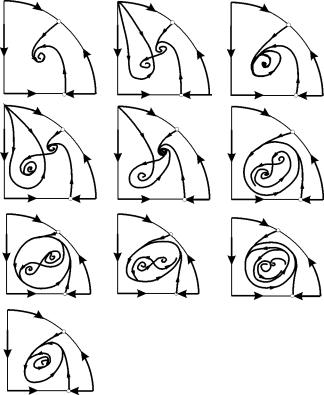
Область притяжения равновесия В 2 (заштрихована) представляет собой “улитку”, скручивающуюся с неустойчивого фокуса В 1 . Если известно, что в начальный момент времени система находилась в окрестности В 1 , то судить о том, придет ли соответствующая траектория в равновесие В 2 или на устойчивый предельный цикл, окружающий три точки равновесия С (седло), В 1 и В 2 можно лишь на основе вероятностных соображений.
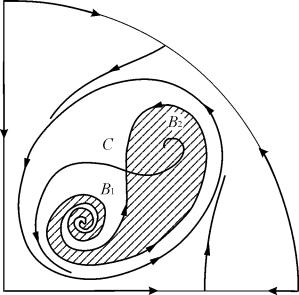
Рис.9.10. Фазовый портрет системы 9.18 для параметрической области 6. Область притяжения В 2 заштрихована
На параметрическом портрете (9.7) имеются 22 различные бифуркационные границы, которые образуют 7 различных типов бифуркаций. Их изучение позволяет выявить возможные типы поведения системы при изменении ее параметров. Например, при переходе из области 1 в область 3 происходит рождение малого предельного цикла, или мягкое рождение автоколебаний вокруг единственного равновесия В. Аналогичное мягкое рождение автоколебаний, но вокруг одного из равновесий, а именно B 1 , происходит при пересечении границы областей 2 и 4. При переходе из области 4 в область 5 устойчивый предельный цикл вокруг точки B 1 “лопается” на петле сепаратрис и единственной притягивающей точкой остается равновесие B 2 и т.д.
Особый интерес для практики представляет, конечно, выработка критериев близости системы к бифуркационным границам. Действительно, биологам хорошо известно свойство “буферности”, или “гибкости”, природных экологических систем. Этими терминами обычно обозначают способность системы как бы поглощать внешние воздействия. Пока интенсивность внешнего воздействия не превышает некоторой критической величины, поведение системы не претерпевает качественных изменений. На фазовой плоскости это соответствует возвращению системы в устойчивое состояние равновесия или на устойчивый предельный цикл, параметры которого не сильно отличаются от первоначального. Когда же интенсивность воздействия превышает допустимую, система “ломается”, переходит в качественно иной режим динамического поведения, например просто вымирает. Это явление соответствует бифуркационному переходу.
Каждый тип бифуркационных переходов имеет свои отличительные особенности, позволяющие судить об опасности такого перехода для экосистемы. Приведем некоторые общие критерии, свидетельствующие о близости опасной границы. Как и в случае одного вида, если при уменьшении численности одного из видов происходит “застревание” системы вблизи неустойчивой седловой точки, что выражается в очень медленном восстановлении численности к начальному значению, значит, система находится вблизи критической границы. Индикатором опасности служит также изменение формы колебаний численностей хищника и жертвы. Если из близких к гармоническим колебания становятся релаксационными, причем амплитуда колебаний увеличивается, это может привести к потере устойчивости системы и вымиранию одного из видов.
Дальнейшее углубление математической теории взаимодействия видов идет по линии детализации структуры самих популяций и учета временных и пространственных факторов.
Литература.
Колмогоров А.Н. Качественное изучение математических моделей динамики популяций. // Пороблемы кибернетики. М ., 1972, Вып .5.
MacArtur R. Graphycal analysis of ecological systems// Division of biology report Perinceton University. 1971
А.Д.Базыкин “Биофизика взаимодействующих популяций”. М., Наука, 1985.
В.Вольтерра: «Математическая теория борьбы за существование». М .. Наука , 1976
Gause G.F. The struggle for existence. Baltimore, 1934.
Еще в 20-х гг. А. Лотка (Lotka), а несколько позднее независимо от него В. Вольтерра предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтерра. В основе модели лежит ряд допущений:
1) популяция жертв в отсутствие хищника растет экспоненциально,
2) пресс хищников тормозит этот рост,
3) смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций);
4) рождаемость хищника зависит от интенсивности потребления жертв.
Мгновенная скорость изменения численности популяции жертв может быть выражена уравнением
dN ж /dt = r 1 N ж - p 1 N ж N х,
где r 1 - удельная мгновенная скорость популяционного роста жертвы, p 1 - константа, связывающая смертность жертв с плотностью хищника, a N ж и N х - плотности соответственно жертвы и хищника.
Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости и постоянной смертности:
dN х /dt = p 2 N ж N х – d 2 N х,
где p 2 - константа, связывающая рождаемость в популяции хищника с плотностью жертв, a d 2 - удельная смертность хищника.
Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т.е. рост числа жертв лимитируется прессом хищников, а рост числа хищников - недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода популяции жертв из-под контроля хищника, хотя на самом деле такое бывает достаточно часто.
Несмотря на всю условность модели Лотки-Вольтерра, она заслуживает внимания уже хотя бы потому, что показывает, как даже такая идеализированная система взаимодействия двух популяций может порождать достаточно сложную динамику их численности. Решение системы этих уравнений позволяет сформулировать условия поддержания постоянной (равновесной) численности каждого из видoв. Популяция жертв сохраняет постоянную численность, если плотность хищника равна r 1 /p 1, а для того чтобы постоянство сохраняла популяция хищника, плотность жертв должна быть равна d 2 /p 2 . Если на графике отложить по оси абсцисс плотность жертв N ж , а по оси ординат - плотность хищника N х , то изоклины, показывающие условие постоянства хищника и жертвы, будут представлять собой две прямые, перпендикулярные друг другу и координатным осям (рис. 6,а). При этом предполагается, что ниже определенной (равной d 2 /p 2) плотности жертв плотность хищника всегда будет уменьшаться, а выше - всегда увеличиваться. Соответственно и плотность жертвы возрастает, если плотность хищника ниже значения, равного r 1 /p 1 , и уменьшается, если, она выше этого значения. Точка пересечения изоклин соответствует условию постоянства численности хищника и жертвы, а другие точки на плоскости этого графика совершают движение по замкнутым траекториям, отражая, таким образом, регулярные колебания численности хищника и жертвы (рис. 6,б). Размах колебаний определяется начальным соотношением плотностей хищника и жертвы. Чем ближе оно к точке пересечения изоклин, тем меньше окружность, описываемая векторами, и соответственно меньше амплитуда колебаний.
Рис. 6. Графическое выражение модели Лотки-Вольтера для системы хищник-жертва.
Одна из первых попыток получения колебаний численности хищника и жертвы в лабораторных экспериментах принадлежала Г.Ф. Гаузе (Gause). Объектами этих экспериментов были инфузория парамеция (Paramecium caudatum ) и хищная инфузория дидиниум (Didinium nasutum ). Пищей для парамеции служила регулярно вносимая в среду взвесь бактерий, а дидиниум питался только парамециями. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности приводил к полному истреблению жертв, после чего вымирала и популяция самого хищника. Усложняя опыты, Гаузе устраивал убежище для жертвы, внося в пробирки с инфузориями немного стеклянной ваты. Среди нитей ваты могли свободно перемещаться парамеции, но не могли дидиниумы. В таком варианте опыта дидиниум съедал всех парамеций, плавающих в свободной от ваты части пробирки, и вымирал, а популяция парамеции затем восстанавливалась за счет размножения особей, уцелевших в убежище. Некоторого подобия колебаний численности хищника и жертвы Гаузе удалось добиться только в том случае, когда он время от времени вносил в культуру и жертву и хищника, имитируя таким образом иммиграцию.
Через 40 лет после работы Гаузе его опыты были повторены Л. Лакинбиялом (Luckinbill), использовавшим в качестве жертвы инфузорию Paramecium aurelia , а в качестве хищника того же Didinium nasutum . Лакинбиллу удалось получить несколько циклов колебаний численности этих популяций, но только в том случае, когда плотность парамеций была лимитирована нехваткой пищи (бактерий), а в культуральную жидкость добавляли метилцеллюлозу - вещество, снижающее скорость движения как хищника, так и жертвы и потому уменьшающее частоту их возможных встреч. Оказалось также, что добиться колебаний хищника и жертвы легче, если увеличить объем экспериментального сосуда, хотя условие пищевого лимитирования жертвы и в этом случае обязательно. Если же к системе сосуществующих в колебательном режиме хищника и жертвы добавляли избыточную пищу, то ответом был, быстрый рост численности жертвы, за которым следовало возрастание численности хищника, приводящее в свою очередь к полному истреблению популяции жертвы.
Модели Лотки и Вольтерры послужили толчком для разработки ряда других более реалистичных моделей системы хищник-жертва. В частности, довольно простая графическая модель, анализирующая соотношение разных изоклин жертвы хищника, была предложена М. Розенцвейгом и Р. Мак-Артуров (Rosenzweig, MacArthur). Согласно этим авторам, стационарная (= постоянная) численность жертвы в координатных осях плотности хищника и жертвы может быть представлена в виде выпуклой изоклины (рис. 7,а). Одна точка пересечения изоклины сосью плотности жертвы соответствует минимальной допустимой плотности жертвы (нижеее популяция подвержена очень большому риску вымирания хотя бы из-за малой частоты встреч самцов и самок), а другая - максимальной, определяемой количеством имеющейся пищи или поведенческими особенностями самой жертвы. Подчеркнем, что речь идет пока о минимальной и максимальной плотностях в отсутствие хищника. При появлении хищника и увеличении его численности минимальная допустимая плотность жертвы, очевидно, должна быть выше, а максимальная - ниже. Каждому значению плотности жертвы должна соответствовать некоторая плотность хищника, при которой достигается постоянство популяции жертвы. Геометрическое место таких точек и есть изоклина жертвы в координатах плотности хищника и жертвы. Векторы, показывающие направление изменения плотности жертвы (ориентированные горизонтально), имеют разную направленность по разные стороны от изоклины (рис. 7,а).

Рис. 7. Изоклины стационарных популяций жертвы (а) и хищника (б).
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 7,б. определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.

Рис. 8. Возникновение колебательных режимов в системе хищник-жертва в зависимости от расположения изоклин хищника и жертвы.
При совмещении изоклин жертвы и хищника на одном, графике возможны три различных варианта (рис. 8). Если изоклина хищника пересекает изоклину жертвы в том месте, где она уже снижается (при высокой плотности жертв), векторы, показывающие изменение численности хищника и жертвы, образуют траекторию, закручивающуюся во внутрь, что соответствует затухающим колебаниям численности жертвы и хищника (рис. 8,а). В том случае, когда изоклина хищника пересекает изоклину жертвы в ее восходящей части (т.е. в области низких значений плотности жертв), векторы образуют раскручивающуюся траекторию, а колебания численности хищника и жертвы происходят соответственно с возрастающей амплитудой (рис. 8,б). Если же изоклина хищника пересекает изоклину жертвы в области ее вершины, то векторы образуют замкнутый круг, а колебания численности жертвы и хищника характеризуются стабильной амплитудой и периодом (рис. 8,в).
Иными словами, затухающие колебания соответствуют ситуации, при которой хищник ощутимо воздействует на популяцию жертв, достигнувшую только очень высокой плотности (близкой к предельной), а колебания возрастающей амплитуды возникают, тогда, когда хищник способен быстро увеличивать свою численность даже при невысокой плотности жертв и таким образом быстро ее уничтожить. В других вариантах своей модели Poзенцвейг и Мак-Артур показали, что стабилизировать колебания хищник-жертва можно, введя «убежище», т.е. предположив, что в области низкой плотности жертв существует область, где численность жертвы растет независимо от количества имеющихся хищников.
Стремление сделать модели более реалистичными путем их усложнения проявилось в работах не только теоретиков, но и экспериментаторов. В частности, интересные результаты были получены Хаффейкером (Huffaker), показавшим возможность сосуществования хищника и жертвы в колебательном режиме на примере мелкого растительноядного клеща Eotetranychus sexmaculatus и нападающего на него хищного клеща Typhlodromus occidentalis . В качестве пищи для растительноядного клеща использовали апельсины, помещенные на подносы с лунками (вроде тех, что используются для хранения и перевозки яиц). В первоначальном варианте на одном подносе было 40 лунок, причем в некоторых из них находились апельсины (частично очищенные от кожуры), а в других - резиновые мячики. Оба вида клещей размножаются партеногенетически очень быстро, и поэтому характер их популяционной динамики можно выявить за сравнительно короткий срок. Поместив на поднос 20 самок растительноядного клеща Хаффейкер наблюдал быстрый рост его популяции, которая стабилизировалась на уровне 5-8 тыс. особей (в расчете на один апельсин). Если к растущей популяции жертвы добавляли несколько особей хищника, то популяция последнего быстро увеличивала свою численность и вымирала, когда все жертвы оказывались съеденными.
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом 6 апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 месяцев, продемонстрировав три полных цикла колебаний численности. Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum ), завезенная в Австралию, значительно снизила численность кактуса опунции, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию.
Говоря о колебаниях хищник-жертва, нельзя не упомянуть и о циклических изменениях численности зайца и рыси в Канаде, прослеженных по материалам статистики заготовок пушнины компанией Гудзон-Бэй с конца XVIII вплоть до начала XX в. Этот пример нередко рассматривался как классическая иллюстрация колебаний хищник-жертва, хотя на самом деле мы видим только следование роста численности популяции хищника (рыси) за ростом численности жертвы (зайца). Что же касается снижения численности зайцев после каждого подъема, то оно не могло объясняться только возросшим прессом хищников, а было связано с другими факторами, по-видимому, прежде всего нехваткой корма в зимний период. К такому выводу пришел, в частности, М. Джилпин (Gilpin), пытавшийся проверить, могут ли быть описаны эти данные классической моделью Лотки-Вольтерры. Результаты проверки показали, что удовлетворительного соответствия модели нет, но как ни странно, оно становилось лучше, если хищника и жертву меняли местами, т.е. трактовали рысь как «жертву», а зайца - как «хищника». Подобная ситуация нашла свое отражение и в шутливом названии статьи («Едят ли зайцы рысей?»), по сути своей очень серьезной и опубликованной в серьезном научном журнале.